DNA FOR HUMAN
ubiquitin واکسین(سه شنبه 87 اردیبهشت 3 ساعت 9:46 صبح )
ubiquitin واکسین
ubiquitin پروتئینی کوچک با حفاظت بالاست که در تمام سلولها ی یوکاریت به طور فراگیر بیان می شود . و ubiquitination اشاره به نوعی تعییر پس ترجمه ای پروتئین دارد که درآن پروتئین با پیوند کوالانسی به یک مونومر ubiquitin باند می شود . مهم ترین وظیفه ی ubiquitin علامت گذاری پروتئین برای اثر پروتئوزوم روی آن وتخریب توسط آن است . در کنار این عملکرد ، ubiquitination کنترل ثبات ، عملکرد و جایگزینی داخل سلولی بسیاری از پروتئین ها را بر عهده دارد .
ubiquitin اولین بار در سال 1975 به عنوان پروتئینی kda 5/8 با عملکرد نا شناخته است که در تمام سلول های زنده بیان می شود. عملکرد اصلی ubiquitin وترکیبات مسیر آبشاری ubiquitination در اواخر دهه ی 1980 شناخته شده است .
سیستم ubiquitination ابتدائا" به عنوان یک سیستم پروتئولیتیک وابسته به ATP است ؛که در عصاره ی سلولی موجود است . یک پلی پپتید مقاوم به گرما ( فاکتور 1 پروتئولیز وابسته به ATP (APF1 ) )یافت شده که چند مولکول از آن به یک مولکول سوبسترای منفرد با پیوند ایزوپپتیدی متصل می شود . این کونژوگه ی سوبسترا – پروتئین به عنوان ubiquitin شناسایی شده است .عملکرد اصلی ubiquitin نشان دار کردن پروتئین ها برای پروتئولیز است.
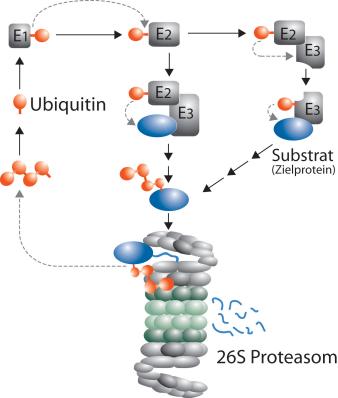
این کونژوگه های APF1 برای تخریب سریع همراه با کاهش APF1 آزاد شناسایی شده اند . پس از اینکه کونژوگه ی APF1 – پروتئین اختصاصی شد، APF1 به عنوان ubiquitin شناسایی می شود . گروه کربوکسیل C ترمینال گلیسین ubiquitin (GLY 76 ) به عنوان بخش کونژوگه شده به رزیدوی لیزین سوبسترا شناخته شده است .
ubiquitin پروتئین کوچکی است که در تمام سلول های یوکاریوتی وجود دارد . عملکرد اصلی معروف نشاندار کردن پروتئین ها به منظور تخریبی به نام پروتئولیز است .حد اقل چهار مولکول ubiquitin برای عمل پروتئوزوم روی سوبسترا ی پروتئینی واتصال به آن برای تخریب لازم است . با این وجود مثال هایی از پروتئین های مورد هدف پروتئوزوم بدون ubiquitin وجود دارد . ubiquitin می تواند پروتئین های عرضی غشا (مثل رسپتور ) را از غشا خارج کند. مثلا" Monoubiquitination برای علامت دار کردن پروتئین های غشایی برای اثر لیزوزوم روی آن است .
ubiquitin دارای 76 اسید آمینه ودارای جرم مولکولیkda 5/8 است . 96% توالی ubiquitin انسان ومخمر مشابه اند . توالی یوبی کوئیتین انسان به صورت زیر است :
MQIFVKTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIFAGKQLEDGRTLSDYNIQKESTLHLVLRLRGG
{kind=link}
) ubiquitylation (ubiquitination.
فرآیند نشانه گذاری پروتئین با ubiquitin (ubiquitination ) شامل یک سری مراحل به شرح زیر است .
1. فعال کردن ubiquitin : فعال شدن ubiquitin طی دو مرحله صورت می گیرد ، که توسط آنزیم فعال کننده ی ubiquitin ، E1 طی فرایند نیازمند به اکسیژن به عنوان منبع انرژی ، صورت می گیرد . مرحله ی اول شاما تولی ماده ی واسطه ی یوبی کوئیتین آدنیلات است . مرحله ی دوم انتقال ubiquitin به جایگاه فعال E1 با مصرف ATP وتولید AMP است . این مرحله در نتیجه ی پیوند تیو استری بین انتهای C ترمینال گروه کربوکسیل ubiquitin وگروه سولفیدریل سیستئین E1 است .
2. انتقال ubiquitin از E1 به سیستئین جایگاه فعال آنزیم کونژوگه کننده ی ubiquitin به نام E2 است که از طریق واکنش ترانس استریفیکاسیون صورت می گیرد .
3. مرحله آخر به طور معمول نیازمند فعالیت یکی از 100 لیگاز های پروتئین - ubiquitin به عنوان E3 (اغلب با اصطلاح لیگاز ubiquitin نام برده می شود . )است . این آنزیم به عنوان محل تشخیص دهنده ی سوبسترای این سیستم است و قابلیت واکنش همزمان با سوبسترا (پروتئین مورد تخریب و E2 را دارد
»» نظرات دیگران ( نظر)
مکانیسم مولکولی اکسین(سه شنبه 87 اردیبهشت 3 ساعت 9:25 صبح )
به نام تدبیر گر اسرار آفرینش
اکسین ها یک کلاس از اجزای رشد گیاهی هستند که اغلب فیتوهورمون یا هورمون گیاهی نامیده می شوند اکسین ها نقش اصلی در تنظیم فرایندهای رفتاری و رشد در چرخه ی زندگی گیاه دارند. نام اکسین از لغت یونانی auxano (رشد کردن) مشتق شده است .آنها اولین هورمون گیاهی اصلی هستند که کشف شدند و اولین سیگنال تنظیمی گیاه هستند. الگوی انتقال فعال درون گیاه پیچیده است. آنها در هماهنگی یا بر خلاف دیگر هورمون ها عمل می کنند. مثلا نسبت اکسین به سیتوکینین در بافت های گیاهی معینی سبب تولید ریشه می شودکه در تضاد با تولید جوانه های بالایی گیاه است.بنابر این یک گیاه می تواند مانند تمام گیاهان بدون نیاز به سیستم عصبی به شرایط محیطی واکنش دهد وبا آنها ارتباط برقرار کند از دیدگاه مولکولی اکسین ها دارای یک حلقه ی آروماتیک و یک گروه کربوکسیلیک هستند.
مهمترین خانواده ی اکسین ایندول3-استیک اسید است که اکثر اثرات اکسین را در گیاهان راه اندازی می کند ومهمترین اکسین طبیعی است. اکسین ها اغلب به علت تحریک ریشه زایی جزء مقاصد اقتصادی مورد استفاده در قلمه زنی و باغبانی هستند. آنها می توانند برای تحریک گلدهی هم شکل وتحریک رسیدن میوه و جلوگیری از افتادن میوه تازه رسیده مورد استفاده قرار گیرند.استفاده ی آنها در دوزهای بالا تولید اتیلن را تحریک می کند. اضافه کردن اتیلن می تواند مانع رشد طولی و سبب افتادن برگ ها وقت افتادن برگ ها شود. برخی اکسین های سنتزی مانند 2-4-D وD5و4-2به عنوان علف کش استفاده می شوند. گیاهان دو لپه ای ببیشترین تاثیر را نسبت به اکسین می پذیرند. نسبت به گیاهان تک لپه مانند علف و کشت های گندمی که تاثیر کمتری می پذیرند. این اکسین های سنتزی عوامل فعال در سیستم عامل یک برگ زدا بودند.
فعالیت های هورمونی
اکسین ها نمو را در همه ی سطوح گیاه تنظیم می کنند از مرحله ی سلولی تا اندام ها و بالاخره تمام گیاه.
مکانیسم مولکولی
اکسین ها مستقیما بیان ژن های ویژه ای را تحریک یا مهار می کنند. اکسین رونویسی را با انتخاب اعضای تخریب گر از پروتئین های مهار کننده ی رونویسی خانواده یAux/IAA القا می کند.عمل تخریب کنندگی AUX/IAA باعث بیان فاکتور واسطه ی رونویسی (ARF) می شود. اکسین های انتخاب شده برای تخریب، با عمل ubiquitination توسط یک لیگاز پروتئینی SCF (کمپلکس چند پروتئینی لیگاز) کاتالیز می شوند. در سال 2005 توضیح داده شد که پروتئین TIR1 F-box که بخشی از کمپلکس لیگاز ubiquitin (SCFTIR1) است یک رسپتور اکسین است. تحت اتصال اکسین به TIR1 مهار کننده های رونویسی ویژه ای برای ubiquitination توسط کمپلکس SCF تخریب می شود. این فرایند نشاندار شده باعث تخریب مهارکننده ها توسط پروتئوزوم و کاستن عمل مهار وهمچنین بیان ژنهای ویژه ای در پاسخ به اکسین می شوند.
پروتئین دیگر به نام ABP1 (پروتئین اتصالی اکسین 1) یک رسپتور است اما نقش آن ناشناخته است. آزمایشات الکترو فیزیولوژیکی با پروتوپلاست ها و آنتی بادی های انتی ABP1 نشان می دهد که ممکن است ABP1 یک عملکرد در غشای پلاسمایی داشته باشد.
در سطح سلولی
در سطح سلولی اکسین برای رشد سلول با تاثیر بر تقسیم و توسعه ی سلولی ضروری است. وابسته به هر بافت خاص اکسین ممکن است رشد محور طولی مانند ساقه های هوایی و توسعه ی جانبی مانند افزایش قطر ریشه یا توسعه ی همه جانبه مانند رشد میوه را تحریک کند. در برخی موارد(رشد کولئوپتیل) توسعه ی سلولی تحت تحریک اکسین در عدم وجود تقسیم سلول رخ می دهد. در دیکر موارد تقسیم سلولی تحت تحریک اکسین و توسعه ی سلولی پس از آن امکان دارد که بلافاصله پشت سر هم درون یک بافت رخ دهد. (ریشه زایی و رشد میوه )در یک گیاه زنده روشن شده که اکسین ها و دیگر هورمون های گیاهی تقریبا همیشه برای تعیین الگوی نمو گیاه باهم دخالت دارند.
طبق فرضیه ی رشد اسیدی فعالیت اکسین اکسین ممکن است مستقیما اولین مرحله رشد طولی سلول راتوسط سلول های مسئول انتقال فعال یون های هیدروژن به بیرون سلول تحریک کند. بنابر این کاهش PH در اطراف سلول ها رخ می دهد. این اسیدیته در منطقه ی دیواره ی سلولی پروتئین های سست کننده ی دیواره معروف به expansins را فعال می کند. که به میکروفیبریل های سلولز اجازه می دهد در دیواره ی سلولی بلغزند و دیواره ی سلولی را به حداقل سختی و غیر قابل انعطافی برسانند. زمانی که دیواره ی سلول با فعالیت اکسین سست شد این دیواره ی با حداقل سستی با فشار تور ژسانس وارد بر دیواره ی سلولی وسیع میشود.
به هر حال فرضیه ی رشد اسیدی افزایش سنتز و انتقال سلول های اولیه دیواره ی سلولی و همچنین فعالیت ترشح دستگاه گلژی را نتوانسته همراه با پدیده ی توسعه ی سلولی توضیح دهد.
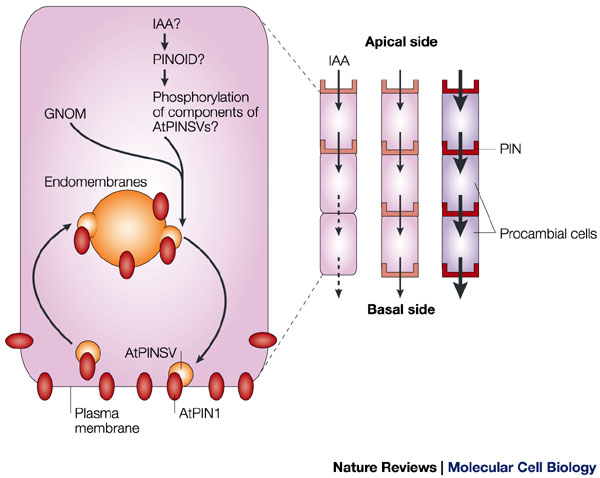
مکانیسم مولکولی انتقال پلار اکسین
حاملین انتقال اکسین به روش انتشار ( PIN پروتئین ها )عامل جریان قطبی اکسین هستند و همچنین مسبب تشکیل ستونهای ممتد در سلولهای پروکامبیومی می باشند . این حاملین از خود سلول بازیافت می شوند و توسط وزیکولهای جدا شده از غشای داخلی به نام اندوزوم به غشای پلاسمایی انتقال می یابند . این حاملین تنها به غشای سلولی در سطح قاعده ای سلول می روند . ودر سطح راسی سلولها این حاملین وجود ندارند .تصور می شود که فاکتور ریبوزیلاسیون گوانین(GNOM ) _گوانین می تواند با آدنین در ADP تبادل شود_ وکیناز سرین – ترئونین (PINOID ) انتقال نامتقارن اندوزوم های حامل پروتئین های PIN را راه اندازی می کنند .
این فرایند با فعال نمودن به ترتیب فاکتور ریبوزیلاسیون ADP وسپس فسفریلاسیون ترکیب ناشناخته ای از اندوزوم ها شروع می شود . راه اندازی تمام این فرآیند با ورود IAA( ایندول استیک اسید ) و ایجاد یک سیکل فید بک مثبت صورت می گیرد .
»» نظرات دیگران ( نظر)
گزرودرما پیگمانتوزوم/2(پنج شنبه 87 فروردین 22 ساعت 11:57 صبح )
پاتوفیزیولوژی
DNAسلولهای سالم ترکیب شده وفرایند ترمیم
پس ازقرار گرفتن آنها درمعرض UV آزمایش شده است.تصحیح نقص ترمیم
پس از برخورد اشعه ی UV به سلول اتصال عرضی ( دیمری شدن )بین بازهای پیریمیدین ایجاد میشود .درسلولهای سالم این بخش از
روش ترمیم حذف نوکلئوتید(NER)
1 . ((GG -NER - NER Global genom
2. TC-NER) ) Tranion Coupled
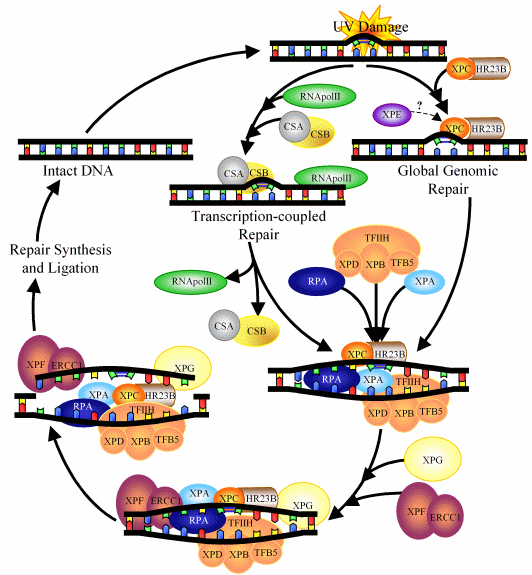
فرآیند نیازمند فعالیت بیش از 30 پروتئین است که به صورت مرحله به مرحله عمل می کند . که این مراحل شامل شناسایی محل آسیب دیده ،باز شدن موضعی
طرح توارثی XP
بیماری XP گروهی از بیماریهای اتوزومی مغلوب است و والدین فرد مبتلا ناقلان ناگزیر یکی از موتاسیون ژنی xp اند .که 25% از فرزندان آنها شانس ابتلا به این بیماری را به شکل همو یگوت دارند و50% هتروزیگوت اند وظاهری سالم دارند اما ناقل این بیماری اند . و25% فرزندان کاملا" سالم هستند . جنسیت در این بیماری هیچ گونه دخالتی ندارد .
»» نظرات دیگران ( نظر)
پروتئین هایPAR نا همزمانی تنظیم کننده های چرخه ی سلولی کیناز شبه(یکشنبه 87 فروردین 11 ساعت 11:39 عصر )
طول چرخه های سلولی به طور وسیعی در بین سلول های مختلف یک جانور متفاوت است . تا به حال مکانیسم تنظیم طول چرخه ی سلولی در حد ناچیزی قابل دست یابی بود . در جنین Caenorhabditis elegans دو سلول با چرخه ی سلولی با مدت زمان متفاوت را تولید می کند که وابسته به عمل پروتئین های قطبی حفاظت شده در برابر بخش بخش شدن آسیب پذیر (PAR ) می با شد . ما نشان دادیم که دو تنظیم کننده ی کلیدی چرخه ی سلولی کیناز شبه چوگان PLK-1 وکیناز فسفاتاز وابسته به سیکلین CDC-25.1 هستند که به طور نا متقارنی در جنین های اولیه پخش شده اند . PLK-1 توسعه ی پیشین سیتوپلاسمی را نشان می دهد . (anterior cytoplasmic enrichment ) و CDC-25.1 توسعه ی وابسته به PLK-1 را در هسته ی پیشین (PLK-1–dependent enrichment in the anterior nucleus) نشان می دهد.هر دو پروتئین برای پیشروی میتوزی نرمال ضروری هستند .
به علاوه این نامتقارنی ها توسط پروتئینهای PAR و پروتئین های افزایش ماهیچه (MEX) MEX-5/MEX-6 که پروتئین دوم به پروتئین کاستن حجم بدن متصل می شود. نتایج ما از مدلی حمایت می کند که به طور نا متقارنی مقادیر PLK-1 را کنترل می کند که به طور نا متقارن CDC-25.1 را برای ایجاد اختلاف در طول چرخه ی سلولی تنظیم کند .
ما پپیشنهاد می کنیم که کنترل PLK-1 و Cdc25 ممکن است در دیگر زمینه های پیشروی مرتبط با تنظیم چرخه ی سلولی باشد .
منبع :
The Journal of Cell Biology, Vol. 180, No. 5, 877-885
»» نظرات دیگران ( نظر)
گزرودرما پیگمنتوزوم( xp )(چهارشنبه 86 فروردین 1 ساعت 12:0 صبح )
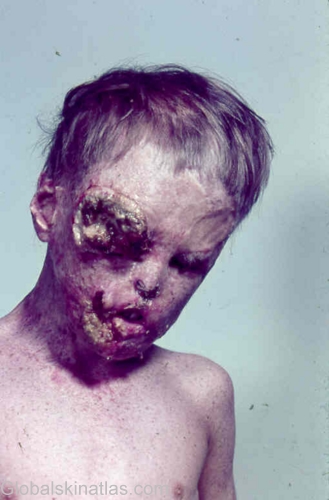
Xeroderma Pigmentosum
گزرودرما پیگمنتوزوم( xp )
اختصاصات بیماری :بیماری xp با اختصاصاتی چون حساسیت به نور خورشید ، درگیر شدن چشم وخطر هزار بربر نئوپلاسمهای پوستی وچشم مشخص شده است .
اغلب افراد مبتلا به xp مستعد گزروزیس (پوست خشک) و پوی کیلودرما (مجموعه ی هیپر- هیپو پیگمانته ، آتروفی واتساع مویرگها ) هستند .سن میانگین ظهور اولیه ی علائم بیماری سرطان غیر ملانومای پوست زیر ده سال است .
پوست : تقریبا" 50% افراد آسیب دیده حساسیت شدیدی به نور خورشید نشان میدهند . که این حساسیت به صورت انتشار آفتاب سوختگی همراه با تاول یا اریتمی در حضور نشر کم نور خورشید است . لکهای ویژه در مناطق در معرض نور خورشید ایجاد می شود؛نوعا" مربوط به بیماری xp است و درکودکان زیر دوسال دیده می شود و به ندرت این نوع لکها درکودکان نرمال دیده می شود .
چشم : ویژگی های غیر طبیعی کننده ی چشم محدود به بخش در معرض اشعه ی uv چشم میشود.قرار گرفتن به صورت ممتد در معرض uv ممکن است باعث انتشار keratitis شود که عامل تیرگی قرنیه وتخلخل آن است .رنگدانه پلکها افزایش می یابد (هیپرپیگمانته )ومژگان تحلیل میروند .
عصب: حدود 30% افراد آسیب دیده علائم عصب شناختی خاصی را بروز میدهند .این علائم شامل میکروسفالی ، نقصان یا فقدان انعکاس های کشیدگی عضلانی تاندون زیرین ، فقدان پیشرونده ی نورون حسی شنوایی و آسیب های مشابه .
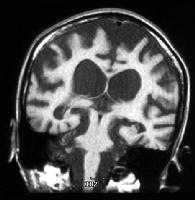
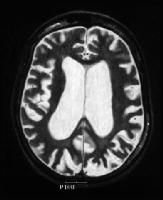
تصویر
»» نظرات دیگران ( نظر)
بازدیدهای امروز: 0 بازدید
بازدیدهای دیروز:0 بازدید
مجموع بازدیدها: 59219 بازدید
