DNA FOR HUMAN
مکانیسم مولکولی اکسین(سه شنبه 87 اردیبهشت 3 ساعت 9:25 صبح )
به نام تدبیر گر اسرار آفرینش
اکسین ها یک کلاس از اجزای رشد گیاهی هستند که اغلب فیتوهورمون یا هورمون گیاهی نامیده می شوند اکسین ها نقش اصلی در تنظیم فرایندهای رفتاری و رشد در چرخه ی زندگی گیاه دارند. نام اکسین از لغت یونانی auxano (رشد کردن) مشتق شده است .آنها اولین هورمون گیاهی اصلی هستند که کشف شدند و اولین سیگنال تنظیمی گیاه هستند. الگوی انتقال فعال درون گیاه پیچیده است. آنها در هماهنگی یا بر خلاف دیگر هورمون ها عمل می کنند. مثلا نسبت اکسین به سیتوکینین در بافت های گیاهی معینی سبب تولید ریشه می شودکه در تضاد با تولید جوانه های بالایی گیاه است.بنابر این یک گیاه می تواند مانند تمام گیاهان بدون نیاز به سیستم عصبی به شرایط محیطی واکنش دهد وبا آنها ارتباط برقرار کند از دیدگاه مولکولی اکسین ها دارای یک حلقه ی آروماتیک و یک گروه کربوکسیلیک هستند.
مهمترین خانواده ی اکسین ایندول3-استیک اسید است که اکثر اثرات اکسین را در گیاهان راه اندازی می کند ومهمترین اکسین طبیعی است. اکسین ها اغلب به علت تحریک ریشه زایی جزء مقاصد اقتصادی مورد استفاده در قلمه زنی و باغبانی هستند. آنها می توانند برای تحریک گلدهی هم شکل وتحریک رسیدن میوه و جلوگیری از افتادن میوه تازه رسیده مورد استفاده قرار گیرند.استفاده ی آنها در دوزهای بالا تولید اتیلن را تحریک می کند. اضافه کردن اتیلن می تواند مانع رشد طولی و سبب افتادن برگ ها وقت افتادن برگ ها شود. برخی اکسین های سنتزی مانند 2-4-D وD5و4-2به عنوان علف کش استفاده می شوند. گیاهان دو لپه ای ببیشترین تاثیر را نسبت به اکسین می پذیرند. نسبت به گیاهان تک لپه مانند علف و کشت های گندمی که تاثیر کمتری می پذیرند. این اکسین های سنتزی عوامل فعال در سیستم عامل یک برگ زدا بودند.
فعالیت های هورمونی
اکسین ها نمو را در همه ی سطوح گیاه تنظیم می کنند از مرحله ی سلولی تا اندام ها و بالاخره تمام گیاه.
مکانیسم مولکولی
اکسین ها مستقیما بیان ژن های ویژه ای را تحریک یا مهار می کنند. اکسین رونویسی را با انتخاب اعضای تخریب گر از پروتئین های مهار کننده ی رونویسی خانواده یAux/IAA القا می کند.عمل تخریب کنندگی AUX/IAA باعث بیان فاکتور واسطه ی رونویسی (ARF) می شود. اکسین های انتخاب شده برای تخریب، با عمل ubiquitination توسط یک لیگاز پروتئینی SCF (کمپلکس چند پروتئینی لیگاز) کاتالیز می شوند. در سال 2005 توضیح داده شد که پروتئین TIR1 F-box که بخشی از کمپلکس لیگاز ubiquitin (SCFTIR1) است یک رسپتور اکسین است. تحت اتصال اکسین به TIR1 مهار کننده های رونویسی ویژه ای برای ubiquitination توسط کمپلکس SCF تخریب می شود. این فرایند نشاندار شده باعث تخریب مهارکننده ها توسط پروتئوزوم و کاستن عمل مهار وهمچنین بیان ژنهای ویژه ای در پاسخ به اکسین می شوند.
پروتئین دیگر به نام ABP1 (پروتئین اتصالی اکسین 1) یک رسپتور است اما نقش آن ناشناخته است. آزمایشات الکترو فیزیولوژیکی با پروتوپلاست ها و آنتی بادی های انتی ABP1 نشان می دهد که ممکن است ABP1 یک عملکرد در غشای پلاسمایی داشته باشد.
در سطح سلولی
در سطح سلولی اکسین برای رشد سلول با تاثیر بر تقسیم و توسعه ی سلولی ضروری است. وابسته به هر بافت خاص اکسین ممکن است رشد محور طولی مانند ساقه های هوایی و توسعه ی جانبی مانند افزایش قطر ریشه یا توسعه ی همه جانبه مانند رشد میوه را تحریک کند. در برخی موارد(رشد کولئوپتیل) توسعه ی سلولی تحت تحریک اکسین در عدم وجود تقسیم سلول رخ می دهد. در دیکر موارد تقسیم سلولی تحت تحریک اکسین و توسعه ی سلولی پس از آن امکان دارد که بلافاصله پشت سر هم درون یک بافت رخ دهد. (ریشه زایی و رشد میوه )در یک گیاه زنده روشن شده که اکسین ها و دیگر هورمون های گیاهی تقریبا همیشه برای تعیین الگوی نمو گیاه باهم دخالت دارند.
طبق فرضیه ی رشد اسیدی فعالیت اکسین اکسین ممکن است مستقیما اولین مرحله رشد طولی سلول راتوسط سلول های مسئول انتقال فعال یون های هیدروژن به بیرون سلول تحریک کند. بنابر این کاهش PH در اطراف سلول ها رخ می دهد. این اسیدیته در منطقه ی دیواره ی سلولی پروتئین های سست کننده ی دیواره معروف به expansins را فعال می کند. که به میکروفیبریل های سلولز اجازه می دهد در دیواره ی سلولی بلغزند و دیواره ی سلولی را به حداقل سختی و غیر قابل انعطافی برسانند. زمانی که دیواره ی سلول با فعالیت اکسین سست شد این دیواره ی با حداقل سستی با فشار تور ژسانس وارد بر دیواره ی سلولی وسیع میشود.
به هر حال فرضیه ی رشد اسیدی افزایش سنتز و انتقال سلول های اولیه دیواره ی سلولی و همچنین فعالیت ترشح دستگاه گلژی را نتوانسته همراه با پدیده ی توسعه ی سلولی توضیح دهد.
مکانیسم مولکولی انتقال پلار اکسین
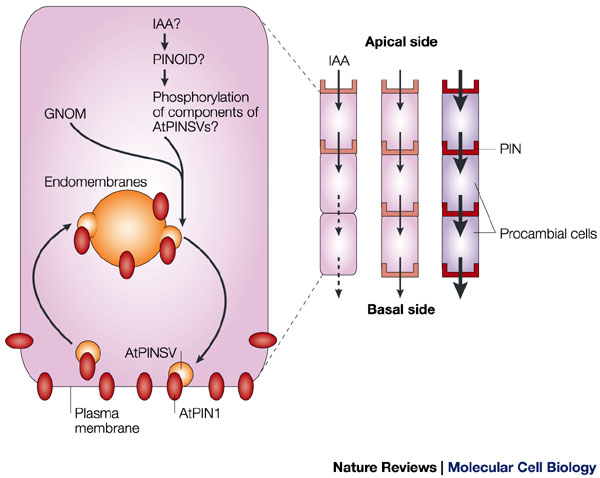
حاملین انتقال اکسین به روش انتشار ( PIN پروتئین ها )عامل جریان قطبی اکسین هستند و همچنین مسبب تشکیل ستونهای ممتد در سلولهای پروکامبیومی می باشند . این حاملین از خود سلول بازیافت می شوند و توسط وزیکولهای جدا شده از غشای داخلی به نام اندوزوم به غشای پلاسمایی انتقال می یابند . این حاملین تنها به غشای سلولی در سطح قاعده ای سلول می روند . ودر سطح راسی سلولها این حاملین وجود ندارند .تصور می شود که فاکتور ریبوزیلاسیون گوانین(GNOM ) _گوانین می تواند با آدنین در ADP تبادل شود_ وکیناز سرین – ترئونین (PINOID ) انتقال نامتقارن اندوزوم های حامل پروتئین های PIN را راه اندازی می کنند .
این فرایند با فعال نمودن به ترتیب فاکتور ریبوزیلاسیون ADP وسپس فسفریلاسیون ترکیب ناشناخته ای از اندوزوم ها شروع می شود . راه اندازی تمام این فرآیند با ورود IAA( ایندول استیک اسید ) و ایجاد یک سیکل فید بک مثبت صورت می گیرد .
»» نظرات دیگران ( نظر)
بازدیدهای امروز: 5 بازدید
بازدیدهای دیروز:2 بازدید
مجموع بازدیدها: 59234 بازدید
